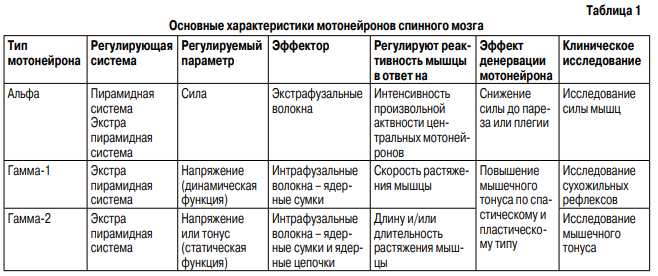
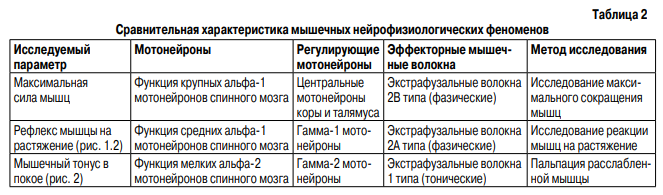
Мышцы обеспечивают весь спектр двигательной активности тела человека. Порой это резкие движения на пределе мощности, в других же случаях требуется очень точное движение с минимальной силой и амплитудой. Сохранение вертикальной позы требует длительного статического напряжения, ходьба и бег – длительной динамической работы мышц. Необходимость нейрофизиологического обеспечения работы мышечной ткани в различных режимах обусловливает наличие нескольких регуляторных систем ЦНС как сегментарного, так и надсегментарного уровня.

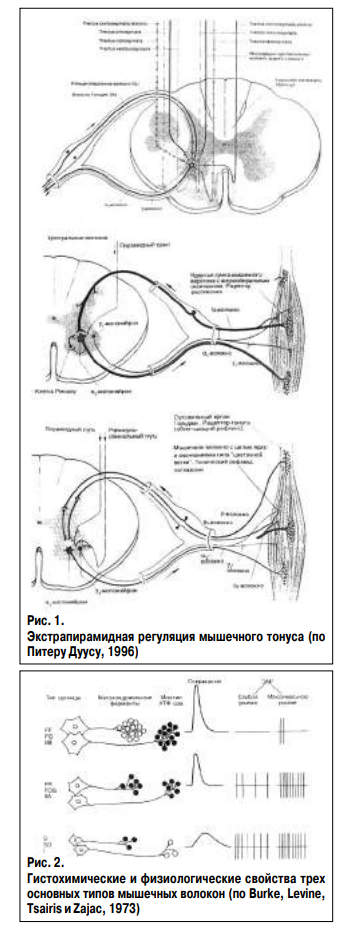
Пирамидная система обеспечивает возникновение мышечного сокращения. Центральные мотонейроны пирамидного тракта оканчиваются на альфа-мотонейронах передних рогов спинного мозга, которые иннервируют экстрафузальные мышечные волокна. Активность альфа-мотонейронов регулируется центральными мотонейронами пирамидного пути (регуляция силы сокращения) и нейронами экстрапирамидных путей (центральная регуляция напряжения) (Дуус П., 1996) – рис. 1. Экстрафузальные мышечные волокна подразделяются на три типа. Волокна 1 (тип S – тонический, аэробный) медленно сокращаются, развивают относительно небольшое напряжение и являются резистентными к утомлению (Мак-Комас А. Дж., 2001). Волокна 1 типа богаты митохондриальными ферментами кислородного дыхания и содержат мало миозиновой АТФ-азы, что определяет способность волокон к длительному сохранению тонуса и неспособность к быстрым интенсивным сокращениям. Имеют развитую капиллярную сеть. Волокна 1 типа отвечают за увеличение и поддержание мышечного тонуса. Волокна 2В типа (тип FF – фазический, анаэробный) характеризуются быстрыми сокращениями, развивают высокое напряжение и быстро утомляются. Волокна содержат небольшое количество митохондриальных ферментов, большое количество миозиновой АТФ-азы и гликогена, имеют слабую капиллярную сеть. Волокна 2В необходимы для обеспечения ударов, бега и прыжков. Волокна 2А типа (тип FR) имеют промежуточные свойства: быструю сократимость, среднее напряжение и резистентны к утомлению, содержат много ферментов кислородного дыхания и много миозиновой АТФ-азы, гликогена. Обладают хорошо развитой капиллярной сетью. Волокна 2А типа обеспечивают быстроту сокращения мышцы и устойчивость ее к утомлению, участвуют во всех видах движений (рис. 2). Каждый тип экстрафузальных волокон иннервируется различными видами альфа-мотонейронов. Волокна 1(S) типа получают иннервацию от мелких, легко возбуждаемых альфа-2 мотонейронов. Аксоны мелких мотонейронов отличаются слабой миелинизацией и медленным проведением импульса. Волокна 2А(FR) типа получают иннервацию от средних альфа-1 мотонейронов. Волокна 2В(FF) типа получают иннервацию от крупных альфа-1 мотонейронов, активируемых только максимальными стимулами. Аксоны крупных альфа-мотонейронов имеют хорошую миелинизацию и приспособлены для быстрого проведения нервного импульса (Зенков Л.Р., Ронкин М.А., 2004). Сила мышечного сокращения регулируется пирамидной системой путем изменения количества двигательных единиц, включившихся в сокращение.
Кроме того, используется механизм рекрутинга альфа-мотонейронов, заключающийся в том, что при сокращении мышцы сначала идет активизация мелких, затем средних альфа-мотонейронов, и только для сокращения с максимальной силой активируются большие альфа-мотонейроны. В отличие от силы, мышечный тонус регулируется экстрапирамидной системой и собственным рецепторным аппаратом мышц: рецепторами нейромышечного веретена и комплекса Гольджи. Тракты экстрапирамидной системы: руброспинальный, тектоспинальный, вестибулоспинальный и ретикулоспинальный заканчиваются не только на альфа-мотонейронах, но и на гамма-мотонейронах передних рогов спинного мозга, которые иннервируют интрафузальные мышечные волокна. Экстрапирамидная система обеспечивает центральный контроль мышечного тонуса. Смысл существования рецепторов нейромышечного веретена состоит в обеспечении обратной связи между мышцей и альфа-мотонейроном. Интрафузальные мышечные волокна и гамма-мотонейроны совместно осуществляют модуляцию обратной связи, изменяя чувствительность рецепторов нейромышечного веретена к
растяжению мышцы и осуществляя тем самым периферический контроль мышечного тонуса (Дуус П., 1996). Выделяют гамма-1 и гамма-2 мотонейроны. Гамма-1 мотонейроны иннервируют интрафузальные волокна с сумками ядер, которые обеспечивают способность мышцы быстро и кратковременно изменять мышечный тонус в ответ на скорость растяжения мышцы. Фактически, чем выше активность гамма-1 мотонейронов, тем активнее вызывается рефлекс мышцы на растяжение и выше сухожильные рефлексы. Гамма-2 мотонейроны иннервируют интрафузальные волокна с сумками и цепочками ядер, которые обеспечивают способность мышцы к постепенному изменению и длительному сохранению мышечного тонуса в ответ на изменение длинны и длительности сокращения мышечного волокна (табл. 1). Клиническое исследование нейрофизиологических феноменов мышечного сокращения включает стандартное измерение силы мышечного сокращения по пятибалльной шкале, а также исследование изменения мышечного тонуса в ответ на растяжение в виде сухожильных рефлексов (рефлекса на растяжение) и мышечного тонуса в покое. Сила мышечного сокращения (функция крупных альфа-мотонейронов) измеряется путем растяжения мышцы при активном максимальном ее сокращении. Динамическая функция мышечного напряжения оценивается с помощью тестирования мышечного рефлекса на растяжение, который вызывается ударом молоточка по сухожилию расслабленной мышцы либо пассивным растяжением мышцы. Дуга рефлекса включает динамические клетки нейромышечного веретена с сумками ядер, афферентное быстропроводящее Iа-волокно и средний альфамотонейрон (альфа-1 мотонейрон).
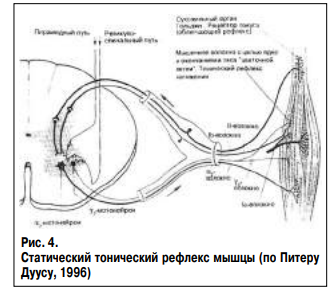
Гамма-1 мотонейрон обеспечивает регуляцию интенсивности рефлекса мышцы на растяжение (рис. 3). В случае повышения мышечного тонуса по спастическому типу расслабленная мышца на растяжение отвечает непроизвольным повышением тонуса, укорочением брюшка и клоническими сокращениями. В случае повышения мышечного тонуса по пластическому типу сокращенная мышца отвечает на растяжение множественным повторением сокращения брюшка по типу миоклонуса, известного как симптом «зубчатого колеса». Статическая функция мышечного напряжения оценивается путем пальпации брюшка расслабленной мышцы (функция мелких альфа-мотонейронов и гамма-2 мотонейронов). Дуга рефлекса включает статические клетки нейромышечного веретена с сумками и цепочками ядер, афферентное Ib-волокно и мелкий альфа-2 мотонейрон. Гамма-2 мотонейрон обеспечивает регуляцию интенсивности тонуса мышцы (рис. 2). Укорочение брюшка встречается при мышечно-тоническом синдроме, повышении мышечного тонуса по спастическому и пластическому типу (табл. 4). Необходимо подчеркнуть, что в любом случае исследуется функция определенного пула альфа мотонейронов.
растяжению мышцы и осуществляя тем самым периферический контроль мышечного тонуса (Дуус П., 1996). Выделяют гамма-1 и гамма-2 мотонейроны. Гамма-1 мотонейроны иннервируют интрафузальные волокна с сумками ядер, которые обеспечивают способность мышцы быстро и кратковременно изменять мышечный тонус в ответ на скорость растяжения мышцы. Фактически, чем выше активность гамма-1 мотонейронов, тем активнее вызывается рефлекс мышцы на растяжение и выше сухожильные рефлексы. Гамма-2 мотонейроны иннервируют интрафузальные волокна с сумками и цепочками ядер, которые обеспечивают способность мышцы к постепенному изменению и длительному сохранению мышечного тонуса в ответ на изменение длинны и длительности сокращения мышечного волокна (табл. 1). Клиническое исследование нейрофизиологических феноменов мышечного сокращения включает стандартное измерение силы мышечного сокращения по пятибалльной шкале, а также исследование изменения мышечного тонуса в ответ на растяжение в виде сухожильных рефлексов (рефлекса на растяжение) и мышечного тонуса в покое. Сила мышечного сокращения (функция крупных альфа-мотонейронов) измеряется путем растяжения мышцы при активном максимальном ее сокращении. Динамическая функция мышечного напряжения оценивается с помощью тестирования мышечного рефлекса на растяжение, который вызывается ударом молоточка по сухожилию расслабленной мышцы либо пассивным растяжением мышцы. Дуга рефлекса включает динамические клетки нейромышечного веретена с сумками ядер, афферентное быстропроводящее Iа-волокно и средний альфамотонейрон (альфа-1 мотонейрон).
Гамма-1 мотонейрон обеспечивает регуляцию интенсивности рефлекса мышцы на растяжение (рис. 3). В случае повышения мышечного тонуса по спастическому типу расслабленная мышца на растяжение отвечает непроизвольным повышением тонуса, укорочением брюшка и клоническими сокращениями. В случае повышения мышечного тонуса по пластическому типу сокращенная мышца отвечает на растяжение множественным повторением сокращения брюшка по типу миоклонуса, известного как симптом «зубчатого колеса». Статическая функция мышечного напряжения оценивается путем пальпации брюшка расслабленной мышцы (функция мелких альфа-мотонейронов и гамма-2 мотонейронов). Дуга рефлекса включает статические клетки нейромышечного веретена с сумками и цепочками ядер, афферентное Ib-волокно и мелкий альфа-2 мотонейрон. Гамма-2 мотонейрон обеспечивает регуляцию интенсивности тонуса мышцы (рис. 2). Укорочение брюшка встречается при мышечно-тоническом синдроме, повышении мышечного тонуса по спастическому и пластическому типу (табл. 4). Необходимо подчеркнуть, что в любом случае исследуется функция определенного пула альфа мотонейронов.
При коррекции дисфункции максимальное усилие сопровождается полным включением двигательных единиц. Органическое поражение нервно-мышечного аппарата исключает возможность восстановления силы ослабленных мышц. Во второй фазе теста исследуется способность мышцы к непроизвольному повышению мышечного тонуса в ответ на длительное (3 секунды и более) изометрическое сокращение мышцы. Данная фаза отражает функциональную активность мелких альфа-мотонейронов и регулирующую функцию гамма-2 мотонейронов. При наличии дисфункции отсутствует увеличение мышечного тонуса и возникает паллидарный тремор. При коррекции дисфункции через 3 секунды после начала изометрического напряжения мышцы возникает усиление напряжения мышцы на 10-15% от исходной величины. В третьей фазе теста исследуется способность мышцы к ответу на резкое растяжение с небольшой амплитудой. Рефлекс мышцы на растяжение отражает функциональную активность средних альфа-мотонейронов и регулирующую функцию гамма-1 мотонейронов. При дисфункции в ответ на быстрое и низкоамплитудное растяжение мышца не отвечает увеличением тонуса. При коррекции дисфункции мышечный ответ на растяжение проявляется в виде усиления напряжения мышцы. Интерференционные кривые мышечного сокращения,
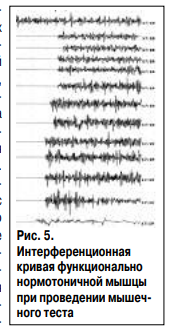
полученные с помощью поверхностной ЭМГ, употребляются для оценки сократительной способности мышц (Николаев С.Г., 2003). Для объективизации выявленных клинических феноменов нейромышечной реактивности использовалось ЭМГ с наложением поверхностных электродов на брюшко мышцы. Произведена запись ЭМГ дельтовидной мышцы пациента А. 62 лет на фоне выполнения мануального мышечного теста (оценка гамма-2 и гамма-1 тонуса). В результате получено 13 интерференционных кривых, отражающих три этапа мышечного теста. Первые 5 кривых отражают исходное напряжение мышцы со средней амплитудой 222-243 мкВ, последующие 5 кривые отражают вторую фазу теста – вазу формирования изометрического напряжения с амплитудой 290-330 мкВ. Следующие 3 кривые отражают третью фазу теста с нарастанием амплитуды до 334-341 мкВ. Визуальное увеличение амплитуды наиболее заметно на предпоследней кривой. Последняя кривая соответствует остаточному тоническому напряжению мышцы после теста с амплитудой 152 мкВ (рис. 5). Дальнейшее исследование проведено в двух режимах: в режиме максимального мышечного сокращения для исследования силы мышцы и в режиме небольшого сокращения мышцы для исследования рефлекса мышцы на растяжение. В каждом режиме исследования регистрировалась ЭМГ кривая в норме и на фоне искусственного подавления нейромышечного аппарата с помощью северного полюса магнита.
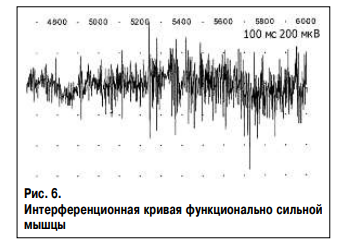
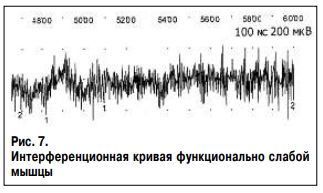
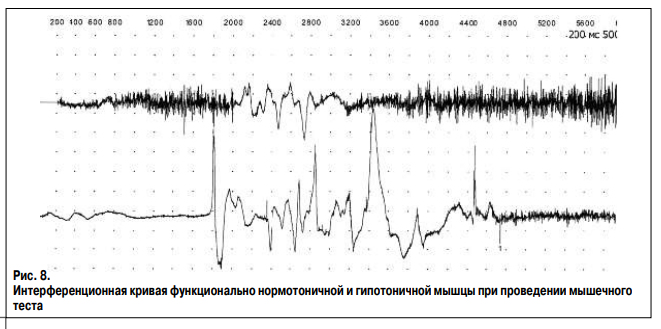
Ниже представлены интерференционные кривые поверхностного ЭМГ исследования общего разгибателя пальцев здорового человека Г., 37 лет без признаков нейромышечной патологии. Исследование проведено на максимальном усилии (рис. 6) и в условиях ослабления мышцы северным полюсом магнита (рис. 7). В результате ЭМГ исследования выявлено снижение амплитуды и частоты нейромышечного ответа. При этом средняя амплитуда колебаний снизилась с 214 до 157 мкВ, средняя частота – с 290 до 156 колебаний в секунду, что свидетельствует об уменьшении числа активируемых нейромыщечных единиц в единицу времени. При исследовании рефлекса мышцы на растяжение записаны две интерференционные кривые с наличием гамма-1 и гамма-2 нормотонии и с наличием гамма-1 и гамма-2 гипотонии (рис. 8). Основными характеристиками интерференционной кривой нормотоничной мышцы (верхняя кривая) являются: 1) короткий период наращивания силы мышечного сокращения без высокоамплитудных колебаний (фазическое сокращение), что свидетельствует об отсутствии паллидарного тремора и высокая амплитуда кривой при последующем тоническом сокращении – гамма-2 нормотония; 2) наличие прироста амплитуды кривой при резком и небольшом растяжении мышцы, что отражает процесс рекрутинга альфа-мотонейронов – гамма-1 нормотония. Основными характеристиками интерференционной кривой гипотоничной мышцы (нижняя кривая) являются: 1) длительный период наращивания силы мышечного сокращения с наличием высокоамплитудных колебаний (фазическое сокращение), что свидетельствует о присутствии паллидарного тремора и низкая амплитуда кривой при последующем тоническом сокращении – гамма-2 гипотония; 2) отсутствие прироста амплитуды кривой при резком и небольшом растяжении мышцы – гамма-1 гипотония.
ВЫВОДЫ :
1.С помощью мануального мышечного теста возможно выделение фазической силы или слабости мышцы, а также гамма-1 и гамма-2 мышечной нормотонии или гипотонии. 2. Интерференционные кривые мышечного сокращения, записанные при поверхностной ЭМГ, позволяют дифференцировать нормотоничный и гипотоничный ответы мышцы по приросту средней амплитуды колебаний.